Article Text

Abstract
Objectives The synovial endothelium targeting peptide (SyETP) CKSTHDRLC has been identified previously and was shown to preferentially localise to synovial xenografts in the human/severe combined immunodeficient (SCID) mouse chimera model of rheumatoid arthritis (RA). The objective of the current work was to generate SyETP-anti-inflammatory-cytokine fusion proteins that would deliver bioactive cytokines specifically to human synovial tissue.
Methods Fusion proteins consisting of human interleukin (IL)-4 linked via a matrix metalloproteinase (MMP)-cleavable sequence to multiple copies of either SyETP or scrambled control peptide were expressed in insect cells, purified by Ni-chelate chromatography and bioactivity tested in vitro. The ability of SyETP to retain bioactive cytokine in synovial but not control skin xenografts in SCID mice was determined by in vivo imaging using nano-single-photon emission computed tomography-computed tomography (nano-SPECT-CT) and measuring signal transducer and activator of transcription 6 (STAT6) phosphorylation in synovial grafts following intravenous administration of the fusion protein.
Results In vitro assays confirmed that IL-4 and the MMP-cleavable sequence were functional. IL-4-SyETP augmented production of IL-1 receptor antagonist (IL-1ra) by fibroblast-like synoviocytes (FLS) stimulated with IL-1β in a dose-dependent manner. In vivo imaging showed that IL-4-SyETP was retained in synovial but not in skin tissue grafts and the period of retention was significantly enhanced through increasing the number of SyETP copies from one to three. Finally, retention correlated with increased bioactivity of the cytokine as quantified by STAT6 phosphorylation in synovial grafts.
Conclusions The present work demonstrates that SyETP specifically delivers fused IL-4 to human rheumatoid synovium transplanted into SCID mice, thus providing a proof of concept for peptide-targeted tissue-specific immunotherapy in RA. This technology is potentially applicable to other biological treatments providing enhanced potency to inflammatory sites and reducing systemic toxicity.
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-commercial License, which permits use, distribution, and reproduction in any medium, provided the original work is properly cited, the use is non commercial and is otherwise in compliance with the license. See: http://creativecommons.org/licenses/by-nc/3.0/ and http://creativecommons.org/licenses/by-nc/3.0/legalcode
Statistics from Altmetric.com
Introduction
Rheumatoid arthritis (RA) is a systemic, inflammatory autoimmune disorder that presents as a symmetric arthritis associated with swelling and pain in multiple joints. Articular inflammation causes activation and proliferation of synovial tissue with hypertrophy of the lining layer, inflammatory cytokine expression, chemokine-mediated recruitment of inflammatory cells and B cell activation with autoantibody production.1–3 Cytokines such as interleukin (IL)-1, tumour necrosis factor (TNF) and IL-6 are found in great abundance.4 These cytokines mediate cartilage and bone degradation by augmenting matrix degrading enzymes such as aggrecanases and matrix metalloproteinases (MMP) and the activation of osteoclasts, which causes bone resorption.
Antagonists to these cytokines now play a fundamental role in the treatment of RA, most notably anti-TNF. These treatments result in clinical benefits for the majority of patients,3 however, 30% to 40% of patients do not respond. In addition, due to systemic immunosuppression, there is a risk of reactivation of latent infections such as tuberculosis. Tissue-specific targeting of existing and novel treatments has the potential to diminish systemic toxicity while increasing drug concentration at the disease site.
An alternative therapeutic method to decrease proinflammatory cytokine expression is to administer anti-inflammatory cytokines, several of which have been shown to be effective in models of RA.4–11 These include interferon (IFN)β,5 IL-106 and IL-4, which has been shown to reduce cartilage destruction and inhibit neoangiogenesis7 ,8 as well as share some of the anti-inflammatory properties of IL-10.4 ,9–11 Clinical trials using IL-4, however, reported a lack of efficacy. The reason for this is not entirely clear but it is strongly believed that the dose used in the trial was suboptimal. It was speculated that the dose necessary to achieve therapeutic efficacy in the synovium would not be tolerated systemically. One approach to overcome this problem is to target the delivery of cytokine to the specific diseased tissue. Linking cytokines to targeting peptides or antibodies has been shown to be effective in preclinical models for the treatment of cancer,12–14 and a tumour neovasculature homing peptide (NGR) conjugated to TNF (NGR-human TNF) is now entering phase III clinical trials as a systemic agent (http://clinicaltrials.gov, trial identifier NCT01098266).
Despite advances in cancer, this therapeutic modality in chronic inflammatory conditions such as RA has so far been relatively unexplored. Neoangiogenesis in RA, similarly to cancer, leads to an enlarged vascular bed and leucocyte infiltration within the synovial tissues and ultimately accelerates disease progression.15 These new vessels are discontinuous, leaky, and present a dysregulated expression of a number of molecules such as integrins, cell surface proteoglycans, proteases and extracellular matrix components as well as endothelial cell growth factor receptors, which are virtually absent or barely detectable in established blood vessels.16 The differences between these new vessels in RA and normal vessels provide a good opportunity for targeted treatment.
Neoangiogenesis is also observed when RA synovial tissue is transplanted into severe combined immunodeficient (SCID) mice. We have used this xenograft model for identification of tissue-specific synovial homing motifs.17 ,18 This has been validated previously by the application of phage display of random peptides19 ,20 and antibody fragment libraries21 to target microvasculature endothelium (MVE) in various tissues.22 ,23 Our laboratory has identified a synovial endothelium targeting peptide (SyETP) that targets endothelial cells within vessels of human inflamed synovial tissue grafted into SCID mice.18 Here, we present evidence that fusion proteins consisting of the SyETP (CKSTHDRLC) fused to the anti-inflammatory cytokine IL-4 led to specific accumulation of the bioactive cytokine in synovial tissue transplanted into SCID mice and that increasing the number of peptide copies increased that period of retention. Specific retention resulted in increased bioactivity as determined by quantification of signal transducer and activator of transcription 6 (STAT6) phosphorylation in synovial compared to control grafts. This study provides proof of concept that homing peptides are a viable means of targeting therapeutics to the MVE of human synovial tissue, opening up a new avenue for translating these findings into novel treatment strategies for patients with inflammatory arthritis.
Methods
Cells and reagents
These are detailed in the supplementary methods.
Human tissue transplantation into SCID animals
Human synovial tissue was obtained from patients undergoing joint replacement surgery. Control human skin was obtained from plastic surgery procedure. Informed consent was obtained prior to the use of these tissues: ethical approval was obtained from the local ethics committee. The criteria for the selection of tissue is described in the supplementary methods. Beige SCID CB-17 mice (5 weeks old; Charles River, Wilmington, Massachusetts, USA) were maintained under sterile conditions in individually ventilated cages. All procedures were carried out in a sterile environment. Synovium and skin were transplanted as previously described.18 , 24
Construction of fusion proteins
Human IL-4 (hIL-4) cDNA was amplified by PCR (from a plasmid kindly provided by DNAX Corp, Palo Alto, CA, USA), using the primers CCCAAGCTTATGGGTCTCACCTCCCAACTGC and ATCTTTTCAGGAATTCGCTCGAACACTTTGAATATTTCTCTC to add HindIII and EcoRI sites to the 5′ and 3′ ends respectively. After digestion with HindIII and partial digestion with EcoRI due to an endogenous EcoRI site, DNA of the appropriate size was purified by gel extraction and inserted into a pcDNA3 vector encoding an MMP-cleavable site25 flanked by EcoRI and NotI restriction sites. Novel DNA sequences encoding one or three copies of a synovial homing peptide18 and a C-terminal His-tag followed by an ApaI restriction site were synthesised by oligonucleotide annealing (see below) and inserted 3′ of the MMP-cleavable sequence. Full-length IL-4-single peptide (IL-4-SP), IL-4-triple peptide (IL-4-TP) and IL-4-triple scrambled (IL-4-TS) cassettes flanked by HindIII and ApaI sites were then inserted into a pFASTBAC1 (Invitrogen, Paisley, Scotland, UK) vector that had been modified by removal of the multiple cloning site at the BamHI and HindIII sites and replacement with annealed oligonucleotides GATCCAAGGTACCACCGCCAAAGCTTACTAAGTTGGGCCCG (forward) and AGCTCGGGCCCAACTTAGTAAGCTTTGGCGGTGGTACCTTG (reverse). Constructs were verified by DNA sequencing.
Targeting specificity of recombinant fusion proteins: imaging by nano-single-photon emission computed tomography-computed tomography (nano-SPECT-CT)
All three fusion proteins were labelled with 125I (see supplementary methods). Mice were injected intravenously with 100 μl of the iodinated construct (100 μg/ml) with a starting activity of approximately 10 MBq. At 0, 40, 90, 180 and 300 min post injection the mice were imaged using a nano-SPECT-CT animal scanner (Bioscan Inc. Washington, DC, USA) as previously described.26 The animals were kept warm and were anaesthetised using 2% isoflurane for the duration of the scan. Helical SPECT images of the transplants were acquired in 20 projections over 30 min using a four-headed camera with 4×9 (1.4 mm) pinhole collimators. CT images were acquired in 180 projections and 1000 ms exposure time using a 45 kVP x-ray source over 3 min. Radionuclide images were reconstructed using HiSPECT (Scivis GmbH) iterative reconstruction software and fused with CT images using proprietary InVivoScope (Bioscan, Washington, DC, USA) software. A three-dimensional volume-of-interest was defined around each graft to calculate the volume and level of activity (MBq) within. Uptake was expressed as activity (MBq) per mm3 tissue. The equipment and methodology used have been previously validated in this model.26
Detection of pSTAT6 in human tissue
Beige SCID CB-17 mice (5 weeks old; Charles River), two per group, were grafted with two pieces of human synovial tissue and two pieces of human skin as previously described.18 ,24 Once the grafts had established, 100 µl of each of the fusion proteins (70 µg/ml) or 100 µl phosphate-buffered saline (PBS) were administered intravenously. As a positive control, an additional group of mice was injected intra-graft with 50 µl of recombinant (r)IL-4 (100 ng/ml). The proteins were allowed to circulate for 45 min. The mice were then killed, the grafts removed and the cytoplasmic fractions were extracted using NE-PER nuclear and cytoplasmic extraction reagents (ThermoScientific, Southend-on Sea, UK). The samples were analysed by western blot analysis for total STAT6 and phosphorylated STAT6 (see supplementary methods).
Statistical analysis
All statistical analysis was performed using SPSS Statistics V.17.0 (SPSS, Chicago, Illinois, USA). One-way analysis of variance (ANOVA) was used when comparing more than two groups (non-parametric data underwent normal transformation). If the groups differed significantly from each other, a Tukey post test was applied to determine where those differences lie. p Values of less than 0.05 were considered significant.
Results
Design, expression and characterisation of peptide-cytokine fusion proteins
Fusion proteins were designed to allow genetic fusion of SyETP (CKSTHDRLC18) to hIL-4. The first fusion protein was constructed by adding a single copy of SyETP to the IL-4 C-terminal (IL-4-SP) (figure 1A(i)). To increase avidity of binding, a second fusion protein was constructed by adding three copies of the SyETP (IL-4-TP) with the SyETPs separated from each other by a rigid spacer peptide27 (figure 1A(ii)). A 6×His tag was added to each fusion protein to enable purification by affinity chromatography (see supplementary methods). In addition, as the inflammatory microenvironment is enriched in MMP,28 an MMP-cleavable sequence25 was inserted between the hIL-4 and synovial homing peptides, with the aim of enabling release of the hIL-4 from the homing peptide at the disease site. This is expected to perform two functions: firstly, enable the hIL-4 to diffuse and interact with target cells physically separate from those cells recognising the SyETP, and secondly, remove any inhibition of hIL-4 activity, which in theory might be conferred by attachment of the SyETP though we have found no evidence of this in our validation experiments in vitro (see later). In addition, as a short peptide, we would expect the proteolytically released SyETP to be degraded rapidly, thus avoiding local saturation of the SyETP binding sites and potentially enabling increased accumulation of the therapeutic payload. A control construct (IL-4-TS), containing three copies of a scrambled peptide (CRKLHTSDC; figure 1A) was also generated (Figure1A(iii)).

Biochemical characterisation of the baculovirus-expressed fusion proteins. (A) Schematic representation of the three fusion proteins consisting of interleukin (IL-4 linked via a matrix metalloproteinase (MMP) cleavage site to (i) one IL-4-single peptide (IL-4-SP), (ii) three IL-4-triple peptide (IL-4-TP) copies of a synovial endothelium targeting peptide (SyETP) or (iii) three copies of a scrambled peptide IL-4-triple scrambled (IL-4-TS). Multiple copies of SyETP or scrambled peptide were connected by helical linkers. Histidine tags were added to each fusion protein for the purpose of purification. (B) To confirm the MMP cleavage site was accessible and susceptible to cleavage, IL-4-SP, IL-4-TP and IL-4-TS were incubated overnight with or without recombinant (r)MMP1 and immunoblotted for (i) IL-4 and (ii) anti-4xhis using appropriate antibodies. (C) IL-4 bioactivity pre/post incubation with MMP1 was determined by assessing the proliferative response of the IL-4 responsive cell line (TF-1) to increasing concentrations of IL-4-TP and IL-4-TS. Proliferation was assessed by measuring ATP accumulation with a luminescence readout. Data represent mean and SD of triplicate wells and are given in arbitrary luminescence units. Titration data are representative of two independent experiments.
Fusion proteins were expressed in insect cells using a baculovirus system and purified by immobilised metal ion affinity chromatography (see supplementary methods). Incubation with MMP1 in vitro confirmed the susceptibility of the proteins to cleavage, shown by reduction in molecular weight (anti-IL-4 antibody) and loss of reactivity with anti-His antibody, which is expected upon loss of the C-terminal peptide containing the 6×His tag (figure 1B). To confirm the IL-4 in the fusion proteins was bioactive, we employed a bioassay in which the TF-1 human erythroleukaemia cell line proliferates in response to IL-4. IL-4-TP and the scrambled control (IL-4-TS) stimulated TF-1 cell proliferation to the same degree (figure 1C). Upon digestion with MMP1 IL-4 bioactivity was increased in both proteins. IL-4 bioactivities of IL-4-TP and IL-4-TS were comparable (EC50s within twofold) pre/post cleavage with MMP1 (figure 1C)).
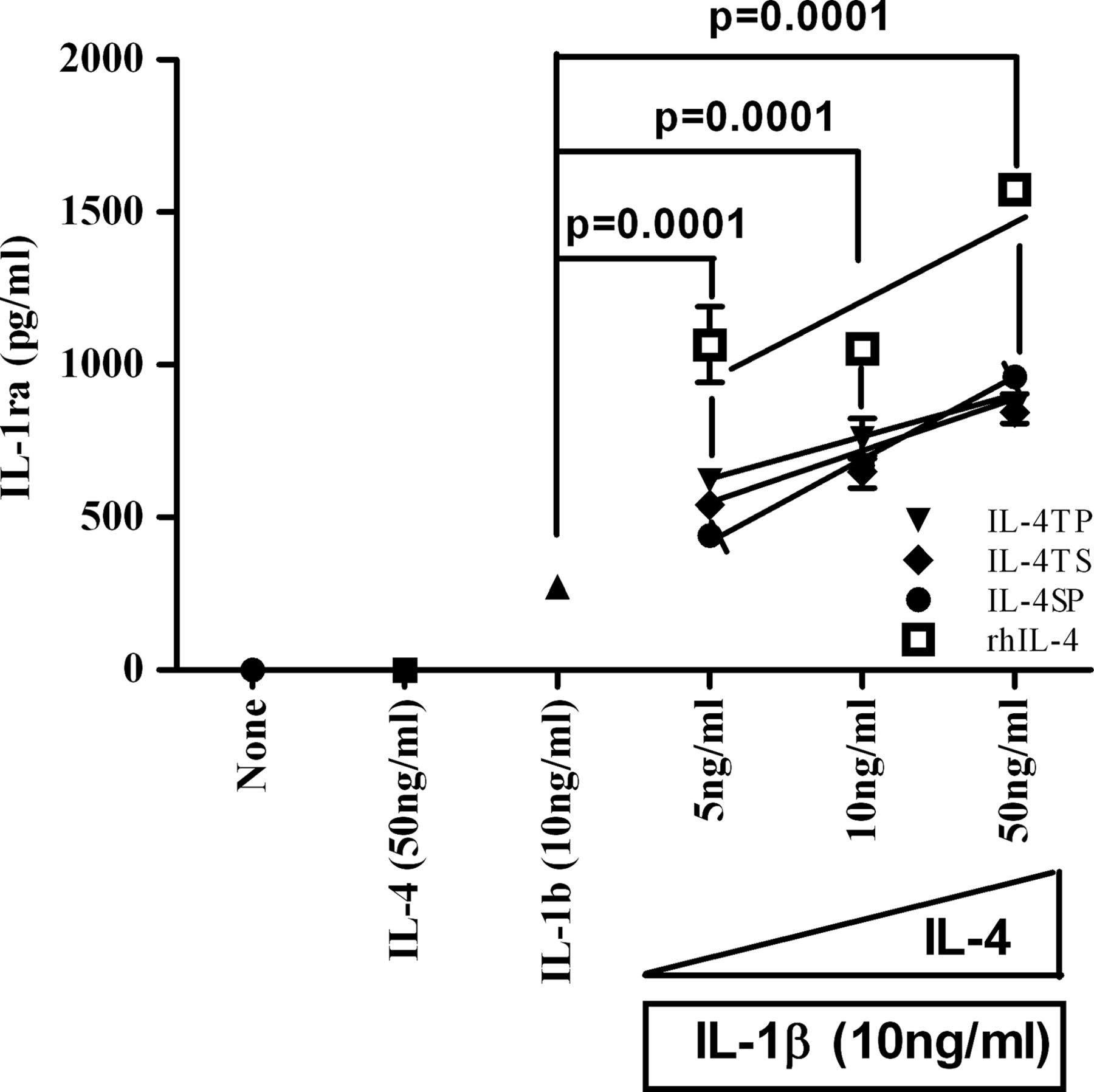
IL-4 synergises with IL-1β to enhance IL-1 receptor antagonist (IL-1ra) production from synoviocytes in vitro
To assess whether SyETP-linked IL-4 retained the capacity of activating anti-inflammatory pathways in synovial cells, primary RA synovial fibroblasts (RASF) and osteoarthritis synovial fibroblasts (OASF) were stimulated with rIL-4 in the presence of the proinflammatory cytokine IL-1β (see supplementary methods). There was no constitutive expression of IL-1ra from resting synoviocytes isolated from patients with RA and OA (figure 2). Upon stimulation with IL-1β (10 ng/ml), the concentration of IL-1ra in the culture supernatant was measured at 300 pg/ml. Such IL-1ra induction was enhanced in a dose dependent fashion with increasing concentrations of recombinant human (rh)IL-4 (5–50 ng/ml) ranging from 1000–1500 pg/ml. Importantly, rhIL-4 alone did not induce IL-1ra production. The synergistic effect of IL-4 and IL-1β on IL-1ra production was observed for all three fusion proteins, albeit to a different degree. Notably, there were no significant differences in the capability of the three fusion proteins to enhance IL-1ra production. In all cases, preincubation with IL-4 induced a highly significant (p<0.001) increase in the production of IL-1ra over IL-1β alone (figure 2). This indicates that SyETP-linked IL-4 retained the capacity of stimulating the production of an anti-inflammatory cytokine in the presence of IL-1β.

Synergistic effect of interleukin (IL)-1β and IL-4 on IL-1 receptor antagonist (IL-1ra) secretion on human synoviocytes. Synoviocytes were incubated for 60 min with or without recombinant human (rh)IL-4 or IL-4-fusion proteins: IL-4-triple peptide, IL-4-triple scrambled or IL-4-single peptide (as described in figure 1A) before adding 10 ng/ml IL-1β and incubated for a further 72 h. The concentration of IL-1ra in culture supernatant was detected by ELISA. Data represent the mean±SE obtained with synoviocytes from three patients. The data is representative of three independent experiments.
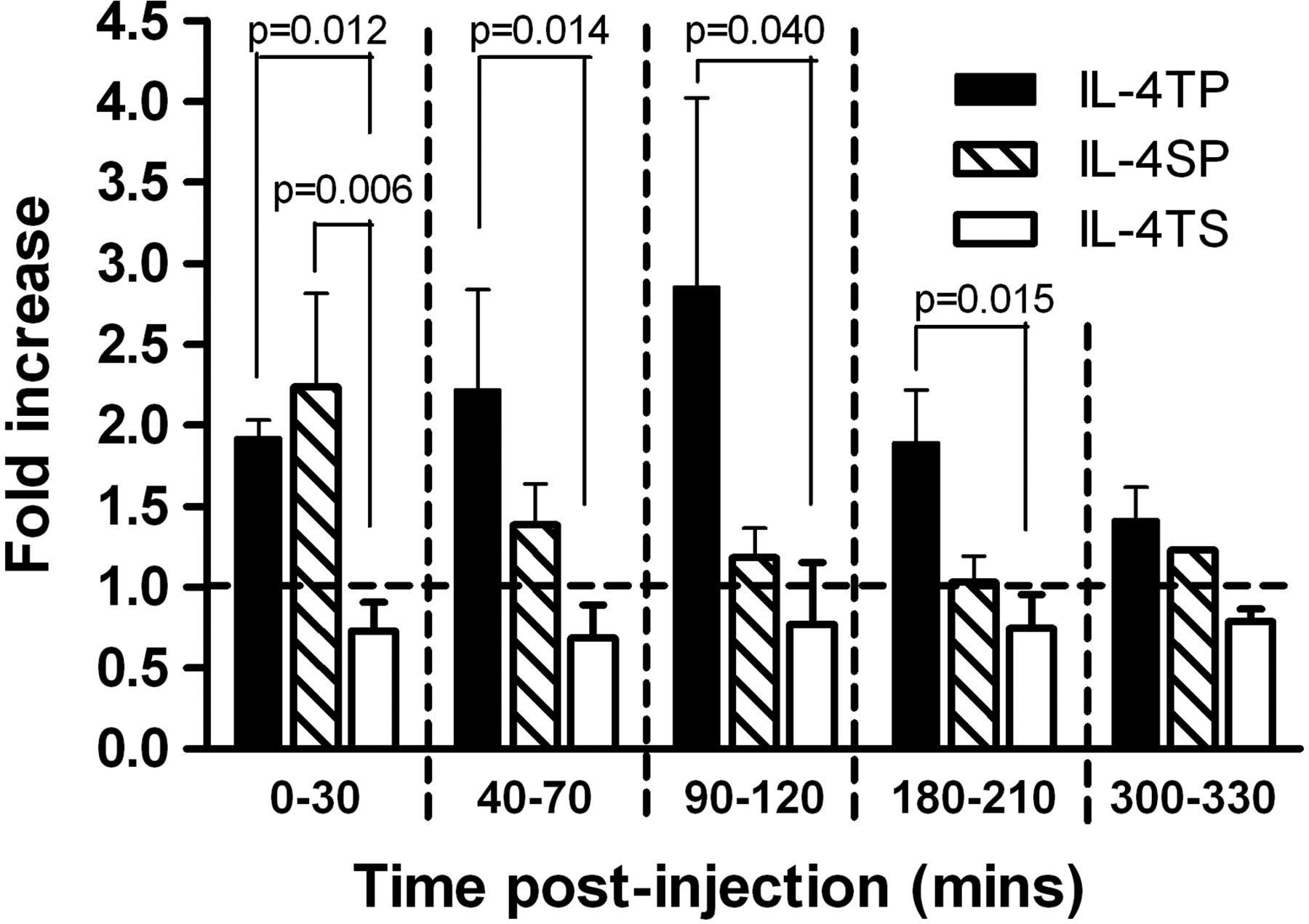
The synovial specific peptide (CKSTHDRLC) retains targeting capability of fusion proteins to synovial grafts in vivo
Having shown in vitro that the IL-4 in the constructs was bioactive and could activate anti-inflammatory pathways, we next wanted to confirm that the SyETP in the fusion proteins had retained synovial targeting capability in vivo. To assess this we radiolabeled the fusion proteins with 125I and administered them intravenously into grafted SCID mice (figure 3). Prior to that experiment we confirmed that radiolabeling did not affect the integrity of the fusion proteins (see supplementary figure 1). In vivo imaging by nano-SPECT-CT allowed quantification of the level of radioactivity in the grafts per mm3 of tissue. To allow direct comparison of the results from individual mice and to normalise for the levels of radioactivity administered, the data are expressed as a ratio of level of radioactivity retained by synovium over that in skin control per mm3 of tissue at each time point. Figure 3 shows a twofold increase in the level of activity in the synovium versus the skin for IL-4-SP during the first 30 min post injection. Importantly, by exploiting the increased avidity of the IL-4-TP the higher activity could be extended to 180 min, peaking between 90–120 min post injection. In contrast, the scrambled control (IL-4-TS) showed no enhanced retention by the synovial graft and dissipated from synovial and skin xenografts at the same rate. These data confirm that SyETP (CKSTHDRLC) leads to preferential accumulation of fusion proteins in synovial compared to control skin grafts.

Preferential accumulation of synovial endothelium targeting peptide-interleukin (IL)-4 constructs in synovial but not in skin xenografts. Severe combined immunodeficient (SCID) mice (seven per group) were grafted with human synovium and skin on either side of the animal subcutaneously in a dorsal position distal to the shoulder joints. A period of 10 days was allowed for the grafts to vascularise. Mice were then injected with iodinated IL-4 fusion proteins and imaged by nano-single-photon emission computed tomography-computed tomography (nano-SPECT-CT) for up to 6 h. The levels of radioactivity per mm3 of tissue in the two grafts were determined by region of interest (ROI) analysis of the images and the ratio between the uptake in synovium and skin transplants calculated. The ratios are shown for IL-4-triple peptide, IL-4-triple scrambled and IL-4-single peptide fusion proteins as described in figure 1A. Data represent mean±SE.
Systemic administration of SyETP-linked, but not scrambled-peptide-linked IL-4, results in functional bioactivity in synovial but not skin grafts in vivo.
In order to assess whether the IL-4 linked to the SyETP and delivered to synovial grafts maintains functional bioactivity in vivo, we measured STAT6 phosphorylation (pSTAT6) in the targeted synovial and skin control grafts. STAT6 is a T helper 2 (Th2)-associated transcription factor that is activated by IL-4 through phosphorylation in the cytoplasm, which is followed by translocation to the nucleus. Thus, we measured the level of STAT6 phosphorylation in the cytoplasmic fractions of the grafts by western blot following intravenous administration of SyETP fusion proteins. As a positive control, rIL-4 was injected intra-graft. Prior to the in vivo experiments, we confirmed the ability of all three fusion proteins to induce STAT6 phosphorylation by stimulating fragments of synovial tissue (organ culture) in vitro (figure 4A). Without stimulation, no phosphorylated STAT6 could be detected in the cytoplasmic fraction of synovial tissue. However, following stimulation with the fusion proteins and rIL-4, phosphorylated STAT6 was clearly detected. The level of phosphorylation was similar with each of the proteins (figure 4A). Next we assessed to what degree functional bioactivity was maintained in vivo. As seen in figure 4B(i,ii), rIL-4 administered intra-graft induced phosphorylation of STAT6 in the synovial and skin grafts, with an average increase of 13-fold and 17-fold over the PBS control. These data confirm that synovial and skin grafted human tissues responded well to rIL-4. Conversely, upon intravenous administration of fusion proteins, only low levels of pSTAT6 were detected in the skin grafts. In contrast, IL-4-SP and IL-4-TP induced an average of 8-fold and 11-fold increases respectively in the synovial grafts, approximately 5-fold greater than the scrambled control peptide (p=0.004 and p=0.001 respectively) (figure 4Bi,ii)).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Synovial endothelium targeting peptide (SyETP)-linked interleukin (IL)-4 maintains functional bioactivity when delivered to synovial grafts in vivo. Severe combined immunodeficient (SCID) mice (two per group) were grafted with two fragments (3–5 mm3) of human synovium (n=4) and two fragments (3–5 mm3) of human skin (n=4) subcutaneously. The mice were left for 10 days to allow the grafts to establish and vascularise. The mice were then injected intravenously with IL-4-single peptide (IL-4-SP), IL-4-triple peptide (IL-4-TP), IL-4-triple scrambled (IL-4-TS) (as described in figure 1A) or phosphate-buffered saline (PBS). An additional control group was represented by mice injected intra-graft with recombinant IL-4 (rIL-4). The mice were killed 45 min post injection, the grafts harvested and the level of phosphorylated signal transducer and activator of transcription 6 (pSTAT6) in the cytoplasmic fractions of the grafts determined by western blot. (A) Immunoblotting for pSTAT6 or total STAT6 in the cytoplasmic fractions of synovial tissue stimulated in vitro by IL-4-SP, IL-4-TP, IL-4-TS (as described in figure 1A) or rIL-4. (B) (i) Immunoblotting for pSTAT6 or total STAT6 in the cytoplasmic fractions from one of four grafts in each group. (B) (ii) Fold increase in the ratio of pSTAT6: total STAT6 over the PBS control. Data represent mean±SE of four grafts in each group from two independent experiments. Statistics are shown for IL-4-SP versus IL-TS versus IL-4-TP.
Discussion
Previously we identified a SyETP and shown it to preferentially locate to the synovial xenografts in a human/SCID chimeric model of RA.18 In this study we constructed fusion proteins consisting of SyETP fused to IL-4 and were able to demonstrate retention and enhanced bioactivity in human synovial tissue transplanted into SCID mice. We also show that multiples of cyclic peptides further increase the period of retention of cytokine in the target tissue, most likely through increased binding avidity, thus providing a strategy for overcoming the general low affinity binding properties of peptides. In addition, specific retention resulted in a significant increase in the bioactivity of IL-4 (STAT6 phosphorylation) in synovial compared to control tissue in vivo, while we confirmed the capacity of these fusion proteins to exert anti-inflammatory activity by measuring the induction of an anti-inflammatory mediator: IL-1ra by primary human RASF in the presence of proinflammatory cytokines in vitro.29 ,30
Application of anti-inflammatory cytokines may be a useful therapeutic strategy in RA, particularly in patients unresponsive to current treatments. However, cytokines are rapidly cleared from the circulation so require high systemic doses to achieve efficacy that normally would prevent their use because of toxicity. One way to overcome these problems is through the use of targeting moieties specific for diseased tissues. To date, preclinical studies targeting cytokine to the inflamed joints in RA have used either rodent or humanised antibody fragments in animal models. Recent studies have also shown that some tissue-targeted short peptides can suppress joint inflammation in the absence of a fusion partner.31 ,32 Our study is the first to demonstrate the use of peptides to target cytokine to human synovial tissue. The transplanted SCID model enables in vivo studies using human disease tissue and therefore rapid in vivo functional analysis of species-specific proteins.33 The fusion proteins constructed in this study have been shown to increase the therapeutic index of the cytokine while retaining the short serum half-life of the cytokine (approximately 30 min, comparable to that of free IL-4 (12–19 min34 ,35)), enabling unbound protein to be rapidly cleared from the circulation, and the capacity to activate anti-inflammatory pathways.
Targeting of cytokine has proved successful in the treatment of cancer in preclinical models12 ,13 ,36 and one such treatment consisting of a NGR conjugated to TNF (NGR-human TNF) is now entering phase III clinical trials as a systemic agent (http://clinicaltrials.gov, trial identifier NCT01098266).37 In contrast, the development of targeted treatments in RA is less advanced and has focused mainly on the development of targeted anti-TNF or anti-TNF-receptor treatment.38–40 Tissue targeting has the potential to enlarge the therapeutic window of the drug it is applied to, thus in the long term may increase the safety and diversity of new and existing therapeutics.
Work is in progress to identify the molecule that binds the target peptide, as this is critical to further develop other targeting moieties and also to improve our understanding at the molecular level of what makes the synovial microvasculature different from that at other tissues or organs. It is hoped that this targeting strategy will improve the therapeutic window for treatment of RA when applied to currently used therapeutics as well as yet untrialled novel conventional and/or biological agents. Moreover, this work further confirms the unique role of the synovium/SCID chimera model for drug development in the context of target validation in human tissues prior to early phase studies in patients.
Acknowledgments
We would like to acknowledge the valuable contribution of Dr Julie Foster and Mr David Ellison for their advice and assistance with labelling and administration of iodinated fusion proteins. We would also like to acknowledge Dr Rebecca Hands for her assistance with the human tissue bank and grading of tissues used in this study and Dr Yarunessa Gofur for contributing to the design of the fusion proteins. In addition our thanks go to Arthritis Research UK for funding this work (grant number: 17606).
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figure 1
- Data supplement 2 - Online supplement
Footnotes
-
Contributors SEW: produced IL-4-fusion proteins, confirmed their ability to activate anti-inflammatory pathways in vitro, designed and performed the in vivo studies and wrote the manuscript. DDiC: built the fusion protein DNA constructs, designed the control protein, produced the fusion proteins, validated their bioactivity in vitro and contributed to the writing of the paper. TEIT: performed one in vivo experiment and the western blot analysis for STAT6 and phospho-STAT6. CMF and SJM: provided technology, technical support and expertise for the in vivo imaging studies. RJ: Provided technical assistance in grafting human tissue onto SCID mice. YKSM: Provided technical advice on the baculovirus system. MB and AN contributed to discussions. YC and CP cosupervised all aspects of the study. SEW and DDiC contributed equally to this work.
-
Competing interests None declared.
-
Patient consent Obtained.
-
Ethics approval East London and The City Research Ethics Committee 3, Floor 2, Room 24 Burdett House, Mile End Hospital, Bancroft Road, E1 4DG.
-
Provenance and peer review Not commissioned; externally peer reviewed.
-
Open Access This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 3.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/3.0/